Nature | 刘颖课题组揭示SAR1B感知细胞内亮氨酸浓度调控mTORC1活性
细胞根据营养状态调控生长是基础的生命科学问题【1】。营养物质包括氨基酸、葡萄糖和脂肪酸等。其中,细胞通过mTORC1复合物感受氨基酸浓度,并据此调节蛋白质合成和细胞生长。氨基酸富足时,mTORC1被活化,启动合成代谢,包括促进游离氨基酸组装成蛋白质,细胞生长加速。相反,游离氨基酸缺乏时,mTORC1被抑制,合成代谢停滞,细胞生长减缓。各种氨基酸的浓度会被其对应的感受器蛋白感知到(也称为氨基酸受体),进而将信号传递给mTORC1【2】。
此外,越来越多的研究表明,mTORC1的持续活化在癌细胞生长和肿瘤发生中扮演重要作用。已发现许多原癌基因和抑癌基因突变通过激活mTORC1促进肿瘤发生。mTORC1抑制剂依维莫司(Everolimus)已被美国食品药品管理局批准用于治疗乳腺癌、胰腺癌和晚期肾癌。另外,还有许多直接或间接的mTORC1抑制剂正在临床试验中【3】。

2021年7月21日,北京大学未来技术学院分子医学研究所、北大-清华生命科学联合中心、北京未来基因诊断高精尖创新中心刘颖课题组在Nature 杂志在线发表题为SAR1B senses leucine levels to regulate mTORC1 signalling的研究论文。报道了一个全新的亮氨酸受体蛋白SAR1B,及其通过mTORC1调控非小细胞肺癌发生的机制。
研究人员结合免疫共沉淀和质谱技术鉴定到亮氨酸受体蛋白SAR1B。SAR1B的经典功能是作为小G蛋白酶参与囊泡形成,并与其他四个亚基共同组装成COPII复合物,在内质网向高尔基体递送物质的过程中发挥重要作用【4】。研究人员发现,SAR1B感知亮氨酸的过程并不依赖于其经典的小G蛋白酶活性。
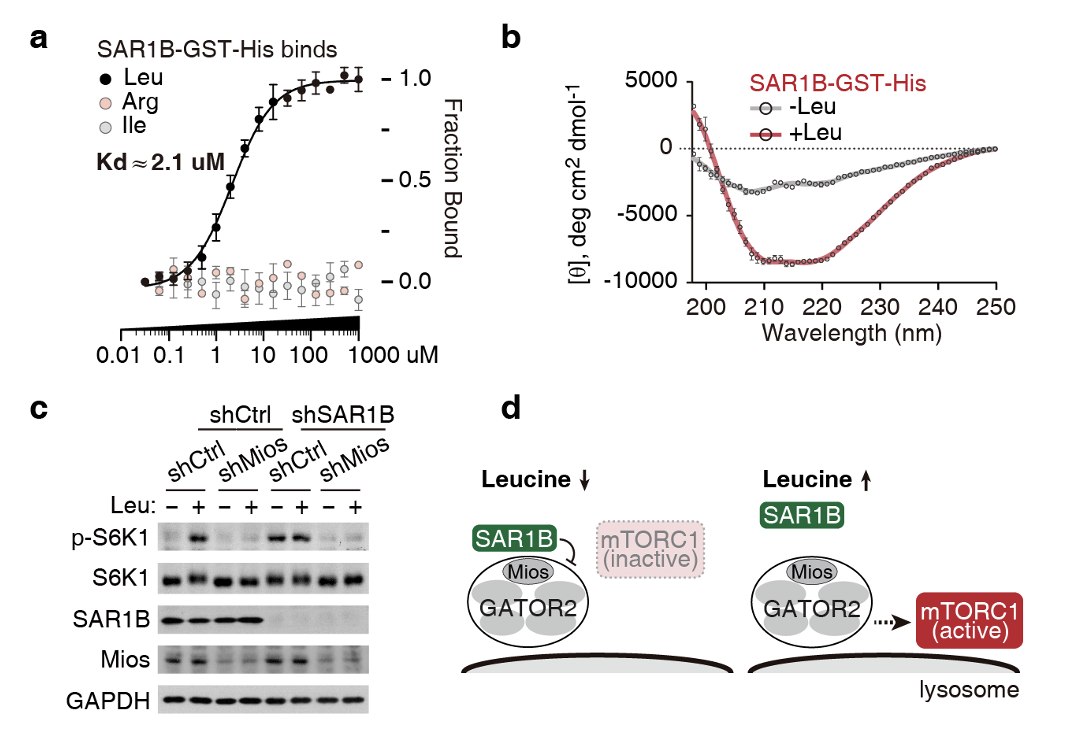
图1. 亮氨酸感知机制。(a) SAR1B体外结合亮氨酸。(b) SAR1B结合亮氨酸变构。(c) SAR1B通过与GATOR2复合物中的Mios蛋白互作调控mTORC1活性。(d)亮氨酸缺乏时,SAR1B抑制GATOR2,mTORC1失活。亮氨酸充足时,SAR1B解离GATOR2,mTORC1被激活。
体外实验表明,SAR1B能够直接结合亮氨酸(图1a),并在结合亮氨酸后发生构象转变(图1b)。细胞实验表明,SAR1B感受到亮氨酸信号后,通过GATOR2复合物将信号传递给mTORC1;敲低GATOR2复合物的组分Mios,将阻断亮氨酸信号的传递(图1c)。
综合诸多实验结果,研究人员总结如下:在亮氨酸缺乏时,SAR1B结合并抑制GATOR2复合物(GATOR2是维持mTORC1活化的重要因子),mTORC1失活。当氨基酸充足时,SAR1B结合亮氨酸,发生变构,随后与GATOR2解离,被解放的GATOR2进而激活mTORC1(图1d)。此外,研究人员还利用无脊椎模式生物线虫,证明了SAR1B感知亮氨酸是进化上高度保守的。 值得一提的是,此前已有一个亮氨酸受体蛋白Sestrin2被报道【5】。研究人员从以下四个方面阐明了这两个受体蛋白的不同之处。(1)它们结合在GATOR2的不同亚基上。SAR1B结合Mios亚基,而Sestrin2结合Seh1L亚基;(2)它们对亮氨酸的敏感程度不同,SAR1B能够感知到更低浓度的亮氨酸,亲和力为Kd ≈ 2 μM。而Sestrin2感知相对高浓度的亮氨酸,亲和力为Kd ≈ 20 μM;(3)它们识别不同的亮氨酸结构特征。SAR1B识别的是亮氨酸的氨基端和侧链,能够容忍羧基端修饰。而Sestrin2则识别亮氨酸的氨基端和羧基端,能够部分容忍侧链修饰;(4)两个受体蛋白存在组织分布差异性,且在肌肉中,两个受体协同调控mTORC1。关于这点,研究人员进行了深入探索。
SAR1B在骨骼肌中丰度最高(图2a),且其表达不依赖于CopII复合物的其他四个亚基(图2b),暗示其存在不依赖于囊泡运输的新功能。骨骼肌中,Sestrin家族另一成员Sestrin1(功能等同Sestrin2)也表达(图2a)。在脂肪组织中,仅Sestrin2高表达(图2a)。功能与受体分布情况一致,肌肉细胞中,SAR1B和Sesrtrin1先后从GATOR2解离,促成了mTORC1 “二次”激活效应(图2c)。而脂肪细胞中,仅Sestrin2对mTORC1进行了“一次”调控(图2 d)。综上,亮氨酸受体SAR1B和Sestrin协作共同调控mTORC1信号(图2 e)。
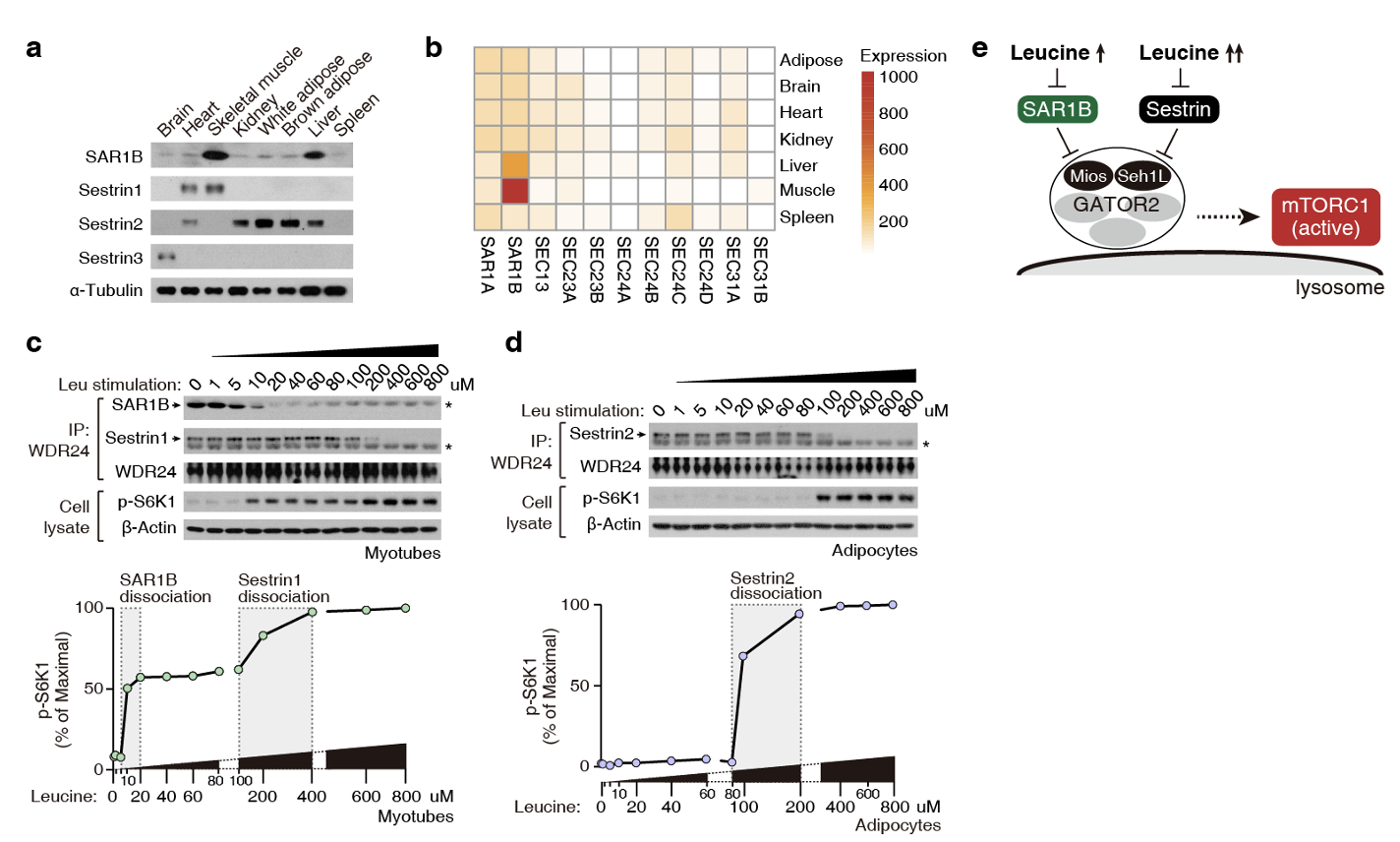
图2. 亮氨酸受体SAR1B和Sestrin协同调控mTORC1。(a) SAR1B和Sestrin1在骨骼肌高表达;Sestrin2在脂肪高表达。(b) SAR1B区别于CopII其他亚基单独在骨骼肌高表达。(c, d) 肌肉和脂肪细胞中亮氨酸感知的差异。(e) SAR1B和Sestrin协同调控mTORC1。
最后,研究人员通过生物信息学分析,发现SAR1B-mTORC1信号在人类非小细胞肺癌中扮演潜在作用,并随后建立了裸鼠肿瘤模型证明了该假设。失去SAR1B及其同源蛋白SAR1A将促进小鼠皮下瘤(图3a, b)和肺内原位瘤(图3c, d)发生,给小鼠注射mTORC1抑制剂雷帕霉素(Rapa.)可阻止肿瘤发生(图3b, d)。此外,肺切片和免疫组化染色结果表明,失去SAR1A/B将促进肿瘤代谢重编程(合成代谢加速,分解代谢抑制),增强肿瘤细胞增殖能力(图3e)。因此,SAR1B在肺癌中发挥着潜在抑癌因子的作用。
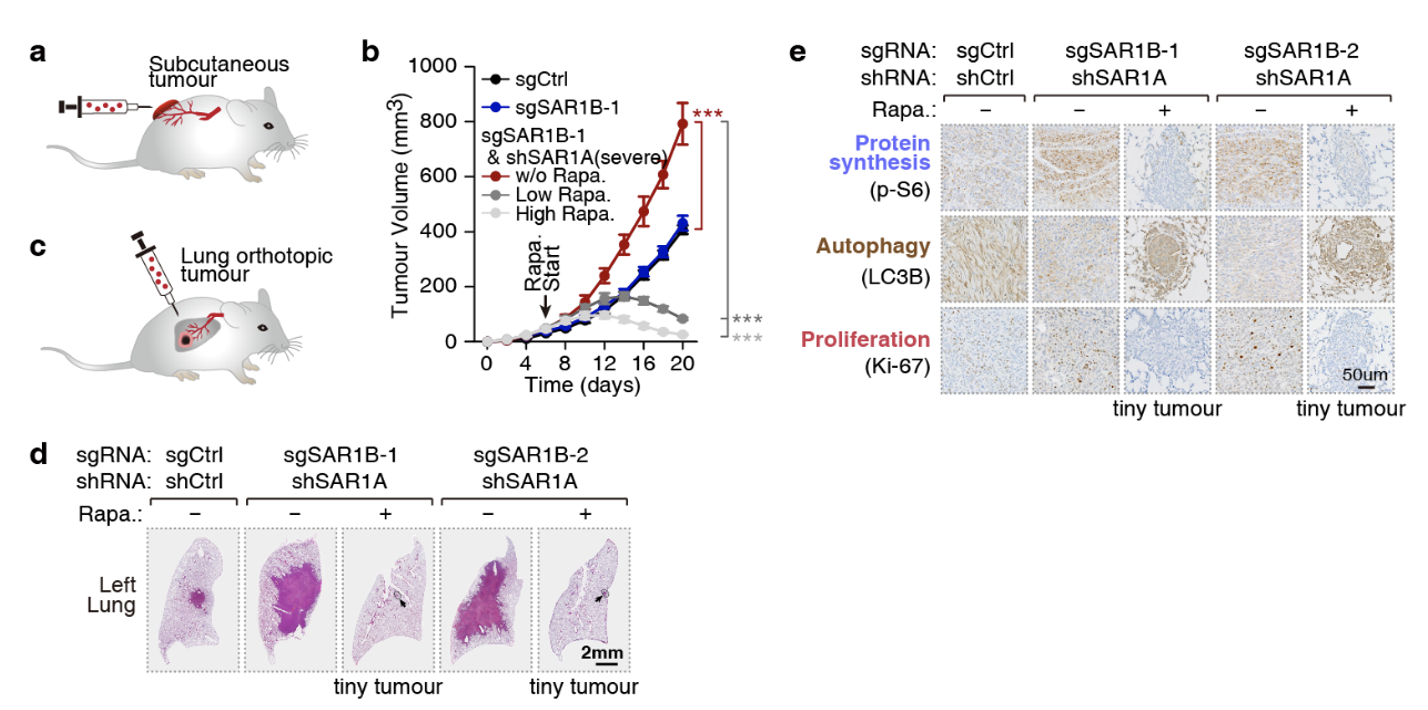
图3. 肿瘤病理意义。(a) 皮下瘤模型。(b) 失去SAR1A/B促进mTORC1依赖的皮下瘤生长。(c)原位瘤模型。(d, e) 失去SAR1A/B促进mTORC1依赖的原位瘤生长(d)和肿瘤代谢重编程(e)。
综上,研究人员发现了一个全新的亮氨酸受体蛋白SAR1B,并详细解析了其感应亮氨酸调控mTORC1信号的分子机制。通过生物信息学分析和建立裸鼠原位瘤模型,研究人员证明了SAR1B是一个潜在的人类非小细胞肺癌抑癌因子,有希望成为非小细胞肺癌的新型生物标志物,为非小细胞肺癌的治疗提供新的药物靶标。
北京大学陈杰博士和刘颖教授为本研究的共同通讯作者。陈杰博士和北京大学未来技术学院分子医学研究所博士生欧宇辉为共同第一作者。该研究获得了国家自然科学基金委、科技部国家重点研发计划、HHMI国际研究学者项目、北京未来基因诊断高精尖创新中心和北大-清华生命科学联合中心的资助。刘颖教授获得了腾讯科学探索奖的支持。
论文链接: https://www.nature.com/articles/s41586-021-03768-w
参考文献:
1.Wullschleger, S., Loewith, R. & Hall, M. N. TOR signaling in growth and metabolism. Cell 124, 471-484, doi:10.1016/j.cell.2006.01.016 (2006).
2.Jewell, J. L., Russell, R. C. & Guan, K. L. Amino acid signalling upstream of mTOR. Nat Rev Mol Cell Biol 14, 133-139, doi:10.1038/nrm3522 (2013).
3.Guertin, D. A. & Sabatini, D. M. Defining the role of mTOR in cancer. Cancer Cell 12, 9-22, doi:10.1016/j.ccr.2007.05.008 (2007).
4.Zanetti, G., Pahuja, K. B., Studer, S., Shim, S. & Schekman, R. COPII and the regulation of protein sorting in mammals. Nat Cell Biol 14, 20-28, doi:10.1038/ncb2390 (2011).
5.Wolfson, R. L. et al. Sestrin2 is a leucine sensor for the mTORC1 pathway. Science 351, 43-48, doi:10.1126/science.aab2674 (2016).